Что может рассказать таз дриопитека о наших предках?
Одна из самых очевидных наших главных особенностей – прямохождение. А оно, разумеется, отражается в строении таза и ноги. Ясно и то, что задолго до спускания с деревьев у предков должны были возникнуть некие преадаптации, облегчившие вставание на две ноги и исключившие другие возможные варианты. А варианты очевидны: павианы слезли с деревьев тогда же и по тем же причинам, что и наши предки, но до сих пор бегают на четвереньках; гориллы и шимпанзе по земле ходят тоже на всех четырёх, да к тому же опираются на землю костяшками согнутых пальцев.
Когда и как закладывались основы нашего необычного способа локомоции – одна из важнейших загадок антропогенеза. В этом направлении сделано уже очень даже немало, но, как обычно в палеонтологии, проблема заключается в редкости и плохой сохранности некоторых костей. Тазовые кости, хотя, казалось бы, очень велики, на самом деле весьма хрупки и быстро разрушаются. Известны считанные единицы тазов наших древнейших предков. А по челюстям с зубами установить тип передвижения как-то не получается.
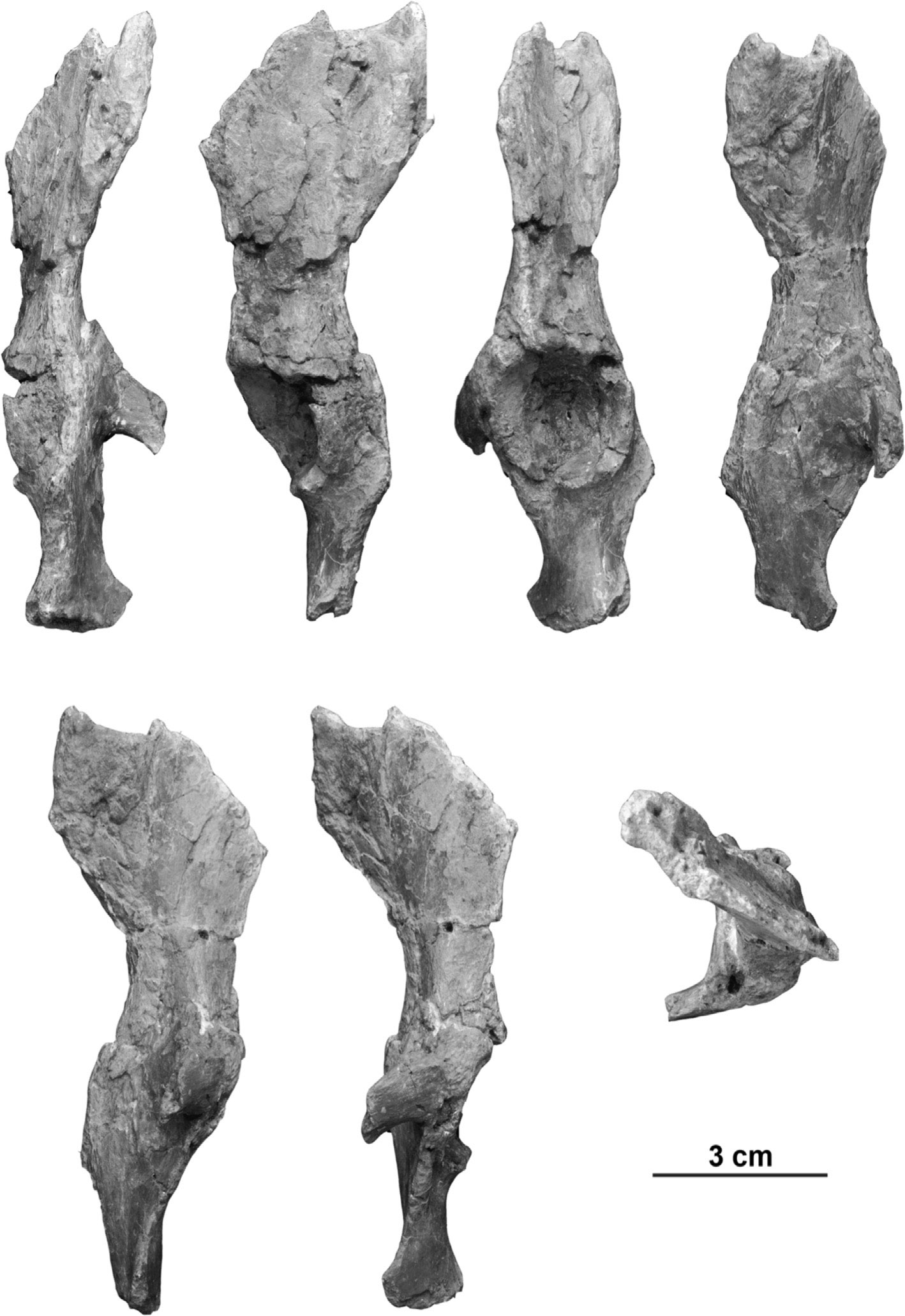
Поэтому важны любые свидетельства, даже фрагментарные. Потому-то особое внимание привлекла находка обломков таза дриопитека из Рудабаньи.
Тут стоит сделать пояснение. Местонахождение Рудабанья находится в Венгрии и датируется временем 9,7 миллиона лет назад (в новой публикации таза дриопитека цифра округлена до 10). Тут обнаружено множество окаменелостей приматов – плиопитеков Anapithecus hernyaki и человекообразных. Последние определялись и как сивапитеки Sivapithecus, они же Ramapithecus hungaricus или Bodvapithecus altipalatus, и как дриопитеки Ataxopithecus serus, Rangwapithecus sericus, Anthropodus brancoi, Dryopithecus brancoi или Hispanopithecus hungaricus, и как самостоятельный род и вид Rudapithecus hungaricus. Авторы статьи – дробители новой волны, а потому использовали именно последнее название. В любом случае важно понимать, что дриопитеки-рудапитеки – не наши прямые предки, это скорее этакие европейские недоорангутаны, своеобразная тупиковая линия, исчезнувшая из-за похолодания и исчезновения тропических европейских лесов (несмотря на школьные учебники и ЕГЭ, провозглашающие дриопитеков нашими пращурами).
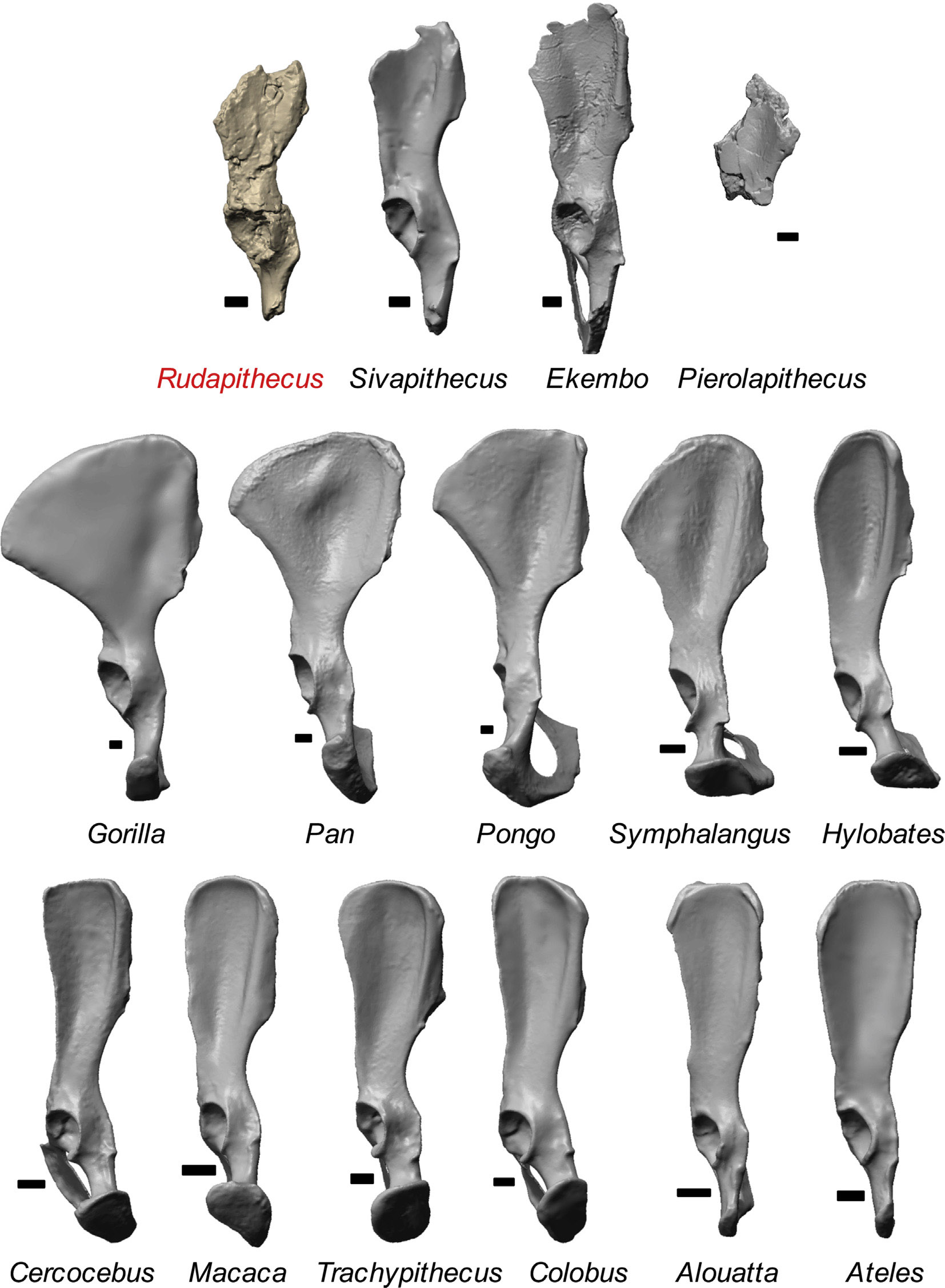
До сих пор в основном палеоприматологам доставались зубы и фрагменты черепов, но теперь повезло ещё больше – нашлись два фрагмента таза: кусок правой вертлужной впадины и седалищной кости и большая часть левой тазовой кости без самого верха подвздошной и переда лобковой. Это даёт возможность изучить многие тонкости, гооврящие о способе передвижения дриопитеков. Сравнительными материалами стали тазовые кости кенийского проконсула (по новомодной традиции называемого в статье Ekembo), индийского сивапитека Sivapithecus, испанского пиеролапитека Pierolapithecus и, конечно, множества современных приматов.
Что же показывает сопоставление всего этого богатства?
В целом, таз дриопитека оказался довольно специфическим, что закономерно и логично, так как речь идёт о самостоятельной эволюционной линии. Но и закономерности показательны.
Вертлужная впадина (ямка сбоку таза, в которую вставляется головка бедренной кости для образования тазобедренного сустава) дриопитека асимметричная, с выступающим передним краем. Такая форма больше всего напоминает версию современных широконосых, макак, гиббонов, сиамангов и орангутанов, но отличается от шимпанзе. Форма крыла подвздошной кости не вполне определима, так как верх не сохранился, но вроде бы больше похожа на вариант мартышкообразных, проконсула, сивапитека и сиаманга; впрочем, это зависит от того, расширялось ли крыло кверху: у мартышек расширения нет, а у человекообразных – есть. Крыло подвздошной кости перпендикулярно лобковой кости, как у гиббонов, в то время как у широконосых, мартышкообразных, проконсула и сивапитека лобковая повёрнута в сторону передних подвзошных остей, а у сиаманга, орангутана, гориллы, шимпанзе и бонобо – задних. Разворот крыла подвздошной вбок сильный, как у сиаманга, орангутана, гориллы, шимпанзе и бонобо, но отличается от слабого у широконосых, мартышкообразных, гиббонов, проконсула и сивапитека, у которых узкая подвздошная идёт вдоль длинного крестца. Тело подвздошной кости дриопитека короткое, как у проконсула, сивапитека и мартышкообразных, на краю распределения более длинной у гиббонов и отличается от очень длинной у орангутанов, горилл и шимпанзе, у коих эта особенность биомеханически связана с укорочением поясницы до 3-4 позвонков. Тело седалищной кости, измеряемое от центра вертлужной впадины до седалищного бугра, короткое, как у гиббонов и орангутанов, чем отличается от длинной у мартышкообразных, проконсула, сивапитека, гориллы и шимпанзе.
Были проанализированы и другие черты, но, думается, нормальному человеку хватит и этого перечисления.
Из всего выше сказанного авторы делают выводы, что таз дриопитека по совокупности черт больше всего похож на версию сиаманга и отличается от мартышкообразных и ранних человекообразных. С этим можно поспорить, из приведённых в статье графиков это ни разу не очевидно: из восьми сопоставлений в трёх случаях действительно есть наибольшее сходство дриопитека с сиамангом (из них в одном – одновременно с сивапитеком, в одном – одновременно с крупными человекообразными, и в одном – одновременно с прочими гиббонами, орангутаном и мартышкообразными), в трёх – с другими гиббонами (из них в двух – ещё и с орангутаном) и в двух – с широконосыми, мартышкообразными, проконсулом и сивапитеком. Выходит, статистика таки не совсем в пользу сиаманга, скорее стоит говорить об общей примитивности с некоторыми чертами специализации. Показательно, что полно совпадений именно с сивапитеком и орангутаном – ближайшими родственниками дриопитеков. Промежуточное положение дриопитека между этими формами как раз логично и закономерно, хотя в статье про это ничего особо не сказано.
В целом, форму таза дриопитека авторы истолковывают как приспособленную к вертикальному лазанию с хватанием за низко расположенные ветки. Такой способ передвижения был характерен для в меру продвинутых гоминоидов, ушедших от исходной древесной четвероногости, но ещё не достигших нирваны развитой брахиации. И именно такой не слишком, но в меру специализированный вариант был важнейшей преадаптацией для нашего прямохождения, не давшей нам спуститься на землю на четвереньках, но и не поставившей слишком жёстких ограничений для развития двуногости.
В очередной раз подтвердилась уже известная истина: многие человеческие черты, отличающие нас от горилл и шимпанзе, вовсе не прогрессивны, а очень даже примитивны. В частности, это касается укорочения седалищной кости и относительного удлинения поясницы. Крупные же человекообразные прошли свой длинный путь эволюции и специализировались по-своему, их нельзя бездумно использовать как модель предка; порой такой моделью выступать должны мы сами.
Источник: Ward C.V., Hammond A.S., Plavcan J.M. et Begun D.R. A late Miocene hominid partial pelvis from Hungary // JHE, 2019.